The third essential organ of your tree is the leaves.
Leaves are the worker bees of the plant world. Tough, tireless little engines, they make almost all of the food that allows the tree to grow itself. Beautifully designed to resist wind, drought, insects and disease, they soldier on until they can endure no more as the killing frost of autumn arrives.
The leaves’ primary task is photosynthesis, using light energy, photons from the sun’s rays. Leaves combine high-energy electrons with CO2 from the atmosphere, and water. The resulting sugars fuel the plant and the world. Without plants fixing carbon into sugars, there is no food base for all life on earth.
Leaves appear from buds on the new shoot either singly (alternating), in pairs (opposite) or in groups (whorled). The leaf can be simple or compound. Compound leaves are made up of small leaflets; there are many types of patterns. Most leaves are flat and bifacial, maximizing surface area. The leaf is attached to the shoot by its stem, the petiole. Outer layers of epidermal cells make up the waxy cuticle, which protects the inner leaf tissue, the mesophyll, from dehydration and predation. The cuticle is present on both upper and lower leaf surfaces, as are the stomata, the leaf’s pores, but there are usually many more stomata on the undersurface, another design aspect to reduce water loss. Leaf water loss is 95% stomatal and 5% cuticular.
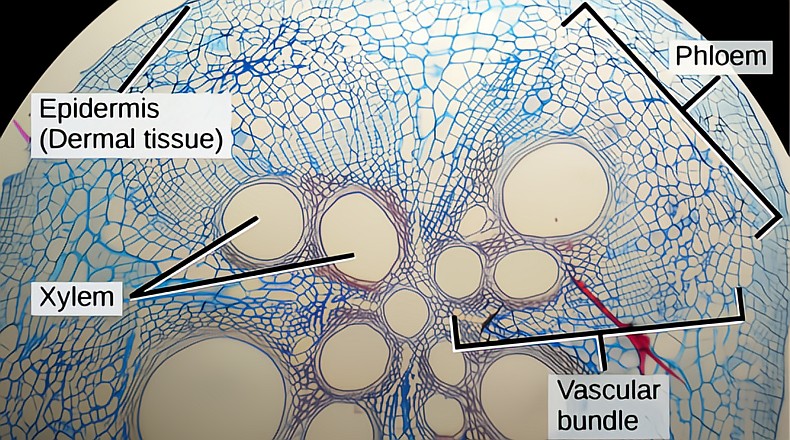
The inner leaf tissue, the mesophyll, is predominantly two types of cells. The upper layer is made up of long palisade cells packed with chloroplasts, the chlorophyll-containing cells that capture light energy for photosynthesis. The lower level is called the spongy mesophyll. Here the cells have a less regular pattern than the palisade. This makes room for the inner leaf air spaces that access the outer atmosphere through the stomata. Braided between these upper and lower tissues are the pipes of the vascular system, the xylem, bringing water and nutrients up, and the phloem, taking away freshly produced sugars made in the chloroplasts. These are the veins we see in leaves.
The vascular system extends throughout the leaf. The xylem/phloem venation is always found in leaves. Parallel venation is found in monocots like a blade of grass. Most dicots or broadleaves have reticulate (spreading) venation, which splits repeatedly into smaller pipes until every cell has contact. Major veins are composed of vascular bundles, or bundle sheaths; a mixture of different types of extra-fortified cells is used, depending on how much strength is needed. The central main vein is the midrib. All sugars produced in the chloroplasts must pass thru the minor vein bundle to be loaded into a phloem sieve tube element.
Light penetrates the surface of the leaf’s epidermis and the cuticle, and moves on into the inner leaf tissue, the mesophyll. Everything about leaf design is to optimize photosynthesis. CO2, another essential component of photosynthesis, only enters the leaf through the stomata. Stomata are the openings, the pores where CO2 can enter the leaf and where water vapour leaves. The CO2 gas has to be in contact with the inner leaf cells, the chloroplasts. The root of that word, chloro, is the same as in chlorophyll; when you see green in plants you see chlorophyll.
Stomata are designed firstly to allow CO2 into the leaf, and secondly, to close when water is limited. This is the enduring dilemma—stay open and allow CO2 in and a lot of water vapor out, or close reducing water loss but shutting down photosynthesis energy production. The stomata openings are circled by two amazing gatekeepers: the guard cells, like a set of lips around the stomata. Guard cells operate by changes in internal water pressure. When water pressure, or turgor, is high in the leaf, the guard cells remain open As water availability drops during the hottest part of a summer day, they close. There is a complicated system of chemical signals between the roots and leaves, and as water availability drops the stomata will close.
An important aspect of leaf/stomata/water interaction is hormonal. During water stress ABA, abscisic acid, increases in leaves, leading to stomatal closing. As K+ potassium ions leave the stomata, there is a drop in turgor, water pressure, and the stomata close.
Where light intensity is low, inside the crown, shade leaves are formed. They are usually larger than sun leaves. Shade leaves are thinner and easier for light to penetrate; their chloroplasts are larger and contain more chlorophyll. Sun leaves have all the light they can use and photosynthesis is only limited by CO2 uptake. Sun leaves have a thinner light-absorbing layer and a thicker CO2-absorbing area (spongy mesophyll). In shade leaves it is the reverse—a thicker light absorbing layer with more chloroplasts and a thinner spongy mesophyll because of the lower need for CO2 uptake.
Xeromorphy is leaf structure that is adapted to persistent dry conditions; it is a hallmark of conifers. The epidermal cells have thick, lignified secondary cell walls and a heavy cuticle. Deeply sunken stomata with a thick waxy layer help prevent water loss. Conifer needle shapes also greatly reduce water loss. The narrow and thin shape reduces the surface to internal volume ratio and subsequent transpiration (water loss) from leaves. Only conifer leaves contain an inner tissue called a fibrovascular bundle. It serves to recover solutes from the transpiration stream and return them to the phloem.
Broadleaves are temporary organs designed for the seasons when it is not freezing. Winter deciduous trees—all of our trees except the conifers—lose their leaves prior to winter in a process called autumnal senescence. This is a two-part event: first the loss of chlorophyll, revealing other pigments in the leaf, and then abscission, which is the programmed removal of plant parts. Senescence is a process of resource recovery; valuable proteins and their amino acids in chloroplasts, synthesized earlier in the year, need to be recovered.
Shorter days and cooler nights signal the start of a process that first changes the color of the leaves and then allows them to drop from the tree. The abscission zone, where the leafs petiole will eventually be removed, starts as a separation zone, and a developing mass of corky cells begins to form at the the base of the petiole. Once mature, this corky abscission zone allows the petiole to split off easily without tearing the twig bark tissue, inhibiting the entry of fungus. Once the petiole is removed, a thin layer of cork cells cover the leaf scar, where the trace of the major veins is still visible.
Timing of leaf emergence next spring is another complicated process. Two aspects of seasonal temperatures affect leaf emergence. Chilling in winter and warm temperatures in spring are needed for the tree to break dormancy. The chilling temperature does not have to be below freezing, though it always is here. Most northern trees given warm conditions in winter before chilling requirements are met are unwilling to begin spring growth. This is good; it prevents trees from starting spring growth during a Chinook in winter. Leaf emergence is also affected by the forcing requirement, a measurement of time above a certain temperature minimum for growth to begin. Most of this is strongly genetically controlled.
Back to Part Two