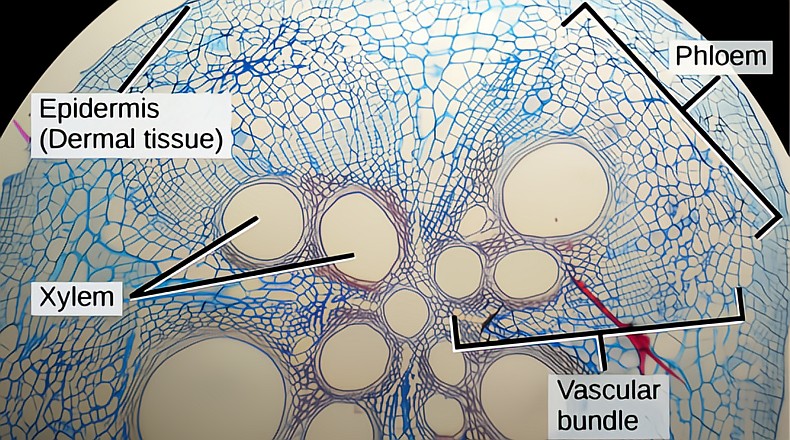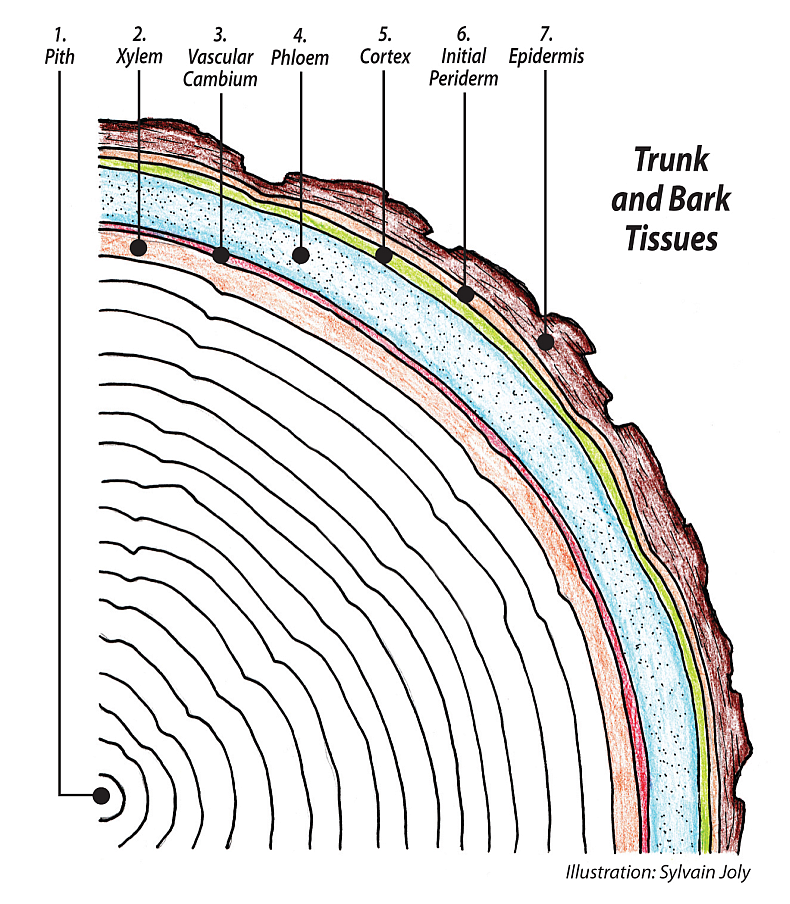
Primary growth accomplished by the SAM and RAM meristems in the shoot’s or root’s first year is how roots and shoots extend in length. Secondary growth, the increase in volume and width of shrub and tree branches and trunks and roots, is accomplished by a lateral meristem called the vascular cambium, VC. The VC wraps continuously around the inner body of the tree under the outer bark, just like your winter base layers cover your own body. The VC is very thin, just a few layers of cells, a bifacial meristem that through a complex ongoing process of mitotic cell divisions grows the whole of the tree trunk. From its outer face it grows all of the tissue of the phloem side, and from its inner face grows all the tissue of the xylem side. The VC is truly the heart, mother, soul of the tree, and is a clear window into the hidden life of trees. Secondary growth allows trees to be what they are, the largest, most numerous and diverse group of land plants.
Whereas there can be thousands of SAMs and RAMs on a tree, there is only one VC running continuously from the roots to the shoots. Knowing on some level that they may live a long time, trees exhibit what can only be called wisdom concerning the functioning of the VC. Imagine how many cell divisions, literally billions, it takes to produce the whole body of a large tree. Mitotic cell division is very complex and intricate; natural mistakes, mutations do occur. This is dangerous in the sense of producing tissue that deviates from the initial genetic plan. The cambium knows this and has come up with an ingenious method to control it. Rather than endless mitotic divisions of the cambium itself, what it does is make copies of itself. These form another outward layer of meristematic cells on each of its faces, both the outer phloem side and the inner xylem side. These are the phloem initials to the outside and the xylem initials to the inside. The VC only needs to produce new initials several times through the entire growing season. This lets the initials do all the heavy lifting, uncounted cell divisions, each year of growing all the new trunk tissues and protects the viability of the VC as it does it. We call the VC and its initials the cambial zone.
The transition of a first year stem’s finished primary growth to one that is capable of producing secondary growth in its second year is a complicated process. The early inner protocambial tissues begin to join and form a mature VC; once this happens it joins the existing VC in older tissue below it. The one-year-old stem is now ready for its second year of life and as the season progresses it adds an additional growth ring produced by its VC.
I am going to follow the VC and its bifacial activities and deal with the phloem side and the xylem side separately. First the phloem side.
The phloem side initials produce two main types of cells. First, the vertically, axially-aligned cells are produced by the fusiform initials. They are the phloem sieve tube elements (STE), their companion cells (CC), and the phloem fibers. These pipes and their supporting fibers run in a vertical direction. The second type are the horizontally aligned ray cells and parenchyma cells produced by the phloem side ray initials. These two types of new cells cross and intermingle like the warp and weft of woven cloth to form the three-dimensional tissue to the outside of the VC. The phloem side fusiform and ray initials produce five main types of cells, the STE, the CC, and the fibers all vertically aligned and the horizontal rays and parenchyma.
The rays themselves have never gotten the attention they deserve. They are the main conduits that pass all the sugars from the phloem required by the VC itself. This movement of sugars continues on deeper inside through the VC and into the xylem parenchyma cells. Live tissues in the xylem, especially storage parenchyma, receive all the sugars and O2 they need from these connected rays. The vessels and fibers of the xylem are dead except for the newest formed vessel tissue which hasn’t lost its inner cell contents yet. Most of the vessels and fibers require no energy because at functioning maturity they are dead. Rays are easy to spot in the end of any crosscut branch or trunk section; they are the spokes of the wheel seeming to radiate outwards from the center.
The Xylem side fusiform and ray initials produce all of the new cells, tissue to the inside of the VC, the greater bulk of the trunk. The vertically-aligned cells of the xylem produced by the fusiform initials are the vessels with perforate ends, perforated with tiny holes to allow water flow, the fibers which are reinforced sclerenchyma cells, and the xylem parenchyma cells. Horizontally-aligned cells produced by the xylem ray initials are the xylem rays and parenchyma cells. The fusiform and ray initials of the xylem side produce five main types of cells to form the xylem tissue. Just like the weaving and intermingling of the three-dimensional phloem tissue the new cells of the xylem side interact to grow the three dimensional tissue we call wood. The xylem side of the new growth accounts for over 90% of the greater annual growth of the trunk; the phloem side is proportionally less.
Gymnosperms (conifers) do things a little differently, being the older first modern pattern of how to be a tree trunk. Instead of the vessels that angiosperms (broadleaf trees) exhibit, conifers have H2O transport cells called tracheids. Longer than vessel elements, they have tapered conical ends that fit well together, strongly overlapping. Tracheid water flow from cell to cell is accomplished by holes called pit pairs. Pits are small holes in the tracheid sidewall and when they line up with the pits of an adjoining tracheid they are called pit pairs. The xylem tissue of conifers consists of three cell types: tracheids, which apart from water conduction function very well as fibers, and parenchyma and rays. On the phloem side of the conifer VC there are also minor differences from their broadleaf cousins. Phloem tissue of conifers consists of sieve cells and albuminous cells, the conifer style of sieve tube elements and their companion cells that we find in broadleaf trees. Other phloem tissue cells in conifer phloem tissue are rays and parenchyma cells. Simpler in design, a little less flashy, the conifer model is one of endurance, proven by 325 million years of adaptation.
Concurrent with the onset of growth in the spring, the cambium is active and already producing the new necessary xylem and phloem tissues. Early wood, the name for the first part of the expansion of the new growth ring, has especially large vessels and tracheids. Plainly seen in any crosscut wood sample, the larger vessel/tracheid cells in their rings are what we use to count the layered growth rings.
Trees have four separate meristems, all providing necessary tissue growth: the SAM and RAM, providing the primary growth of shoots and roots, the VC described above and the periderm or cork cambium providing the secondary growth. The periderm is the name for three collected tissues that always grow together in the rings that form the outer bark. They are, from the inside out, the phelloderm, the phellogen and the phellum or cork. An annually generated tissue, the phellogen, produces all three. Formed anew each year, the phellogen gets its meristematic ability from connected ray parenchyma cells that are genetically programmed to produce the phellogen. The phellogen encircles the cortex to the inside and produces the outer cork, the bark. A new phellogen forms each year from meristematic ray parenchyma cells.