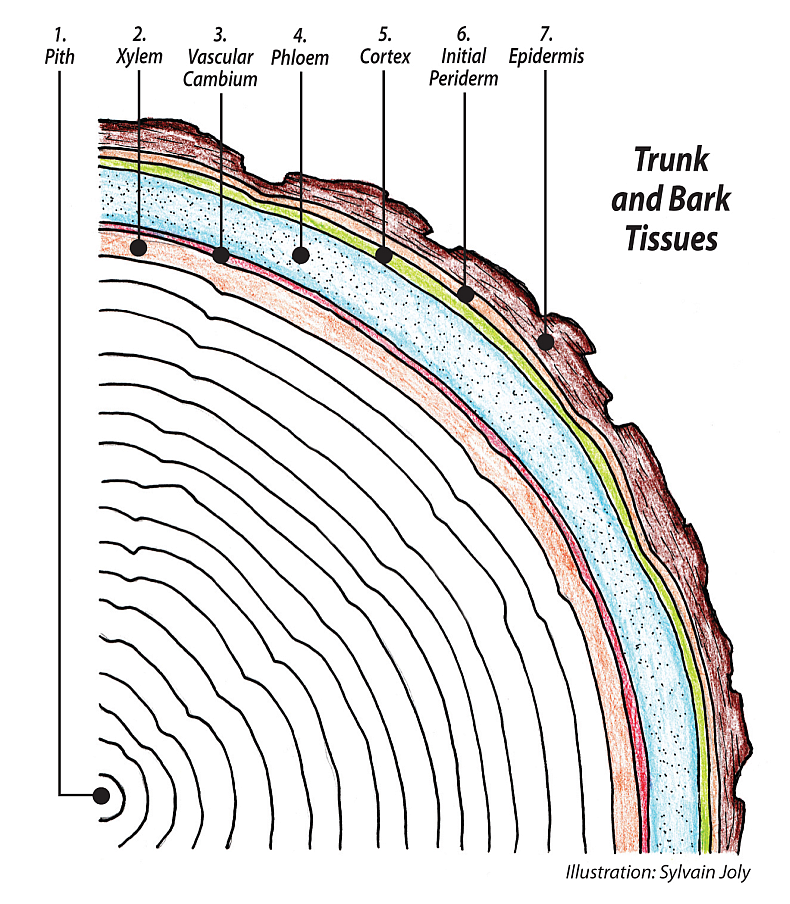
When we talk of meristems we are focused on the processes of primary and secondary growth. Primary growth in trees is the annual extension of shoot and root tips to make the plant taller and expand its root system to match the greater mass of leaves above. Secondary growth is the expansion of the trunk and bark accomplished by the vascular cambium and the cork cambium. Herbaceous plants, annuals, biennials and perennials experience only primary growth, whereas woody plants, trees and shrubs experience both primary and secondary growth. Secondary growth and the vascular and the cork cambiums will be dealt with separately.
The only way that plants can grow and get larger is to increase their mass by making more cells. That is accomplished through a process called mitosis. Mitosis takes us back to the cell cycle and the generation of two daughter cells from a mother cell. This dividing action is what meristems are all about, and the position and activity are strongly genetically controlled. The two meristems that perform primary growth are the RAM (root apical meristem) and the SAM (shoot apical meristem). The RAM is located at the root tips and the SAM is located at shoot tips. The SAM produces all the above-ground organs, stems, leaves and flowers. The RAM produces the root system. Every shoot tip has a SAM and every root tip has a RAM.
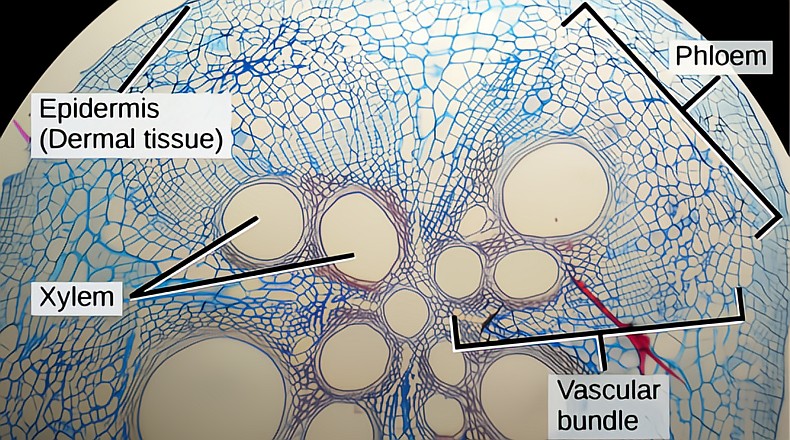
The SAM produces three other specialized meristems that each grow an essential part of the expanding shoot tip: the protoderm, the procambium and the ground meristem.
The protoderm (first skin) generates the outer layer of cells that covers the young expanding leaves. After cell expansion is finished, the protoderm matures into epidermal pavement cells, guard cells and trichomes (leaf hairs). (See Nanking cherry.) The procambium generates the cells that will differentiate into the xylem and phloem strands. The ground meristem generates the ground tissue, the cortex and pith in stems and the mesophyll in leaves. This miracle growth system, gene expression, is run by information stored in the cell’s nucleus and orchestrated by proteins carrying out those instructions.
Besides growing the three meristems that produce primary growth of the epidermis (protoderm), ground meristem (pith and cortex), and the vasculature (procambium), the SAM also grows all the exterior lateral organs—the leaves, axillary buds, flowers and the lateral branches. These lateral external organs begin from the surface of the SAM tip. Shortly after initial growth, the axillary buds may develop their own SAM and continue again as described. Producing their own protoderm, procambium and ground meristem and leaf primordia. Actively expanding growth regions produce the plant hormone IAA (indole acetic acid or auxin), which inhibits growth of axillary buds. This process is called apical dominance.
A common physical example of apical dominance is seen by watching the tops of spruce trees and other conifers. These simple and beautiful patterns are very easy to observe. When we see a spruce tree with a single stemmed leader, the single finger at the top, what we are seeing is undisturbed natural growth. The leader shoot has grown more than any other shoot on any other branch of the tree, and if this growth pattern is undisturbed, then the sharp arrow-headed or conical form of the tree will persist.
The side, or lateral, branches are suppressed by the main leader. The leader also produces the hormone auxin, which inhibits the growth of side shoots. If the leader is removed, pruned off or killed by insect feeding (white pine weevil), a dilemma occurs. Who is the leader? Who will continue the pattern? With spruce trees, sometimes as many as five or six side branches will answer the call. And the quick competitive growth won’t stop until one leader rises above the other, or in many cases, we now have a multi-trunked tree.
Now we need to get back to the RAM, the root apical meristem, and its processes and tissues that grow the root system. The primary growth in length of the roots is accomplished by the RAM where mitotic cell division occurs. The greatest concentration of active cell division occurs at the root tip in the quiescent centre. The quiescent centre is the tissue that, through mitotic division, supplies the mother cells that will divide and divide to produce the new cells needed for root growth.
Just as with the growth we have seen in the SAM, the RAM acts in a similar fashion and produces three types of meristematic tissue that will grow the root. The protoderm produces the epidermis, the procambium produces the vasculature and the ground meristem produces the cortex; there is no pith in roots. All of this ongoing cell division is happening in the area we call the zone of division, which only occurs in roots.
The zones of division, elongation and maturation are the three main growth and expansion areas in the newly developing root. Cells are dividing in the zone of division, the zone of elongation extends the length of the root through the soil and the zone of maturation, which does not extend farther through the soil, is where lateral roots will eventually develop. This genius system never pushes the new lateral roots forward into the soil through rough and sharp particles which would tear them off. Instead the root waits to produce new lateral roots and only does so in the zone of maturation, where they will succeed.
Unlike shoots, the roots only produce one lateral organ: new roots. These lateral roots, unlike the root tip, are not initially produced by the RAM. Lateral roots form internally, back from the root tip, and as they extend outward through existing root tissue, they form their own RAM, and head off in search of water and minerals.
The vasculature, the xylem and phloem in a growing root, are contained in a central core called the vascular stele. Lateral roots are generated by mitotic activity of cells at the outer edge of the stele. The developing lateral root tip pushes through the cortex and epidermis and out into the soil. Meristematic cells in the developing lateral root form its xylem/phloem tissue and connect to the existing vasculature in the stele. A root cap develops at the tip of the new lateral root with its own RAM, and the endlessly spreading and exploring life of the roots goes on.