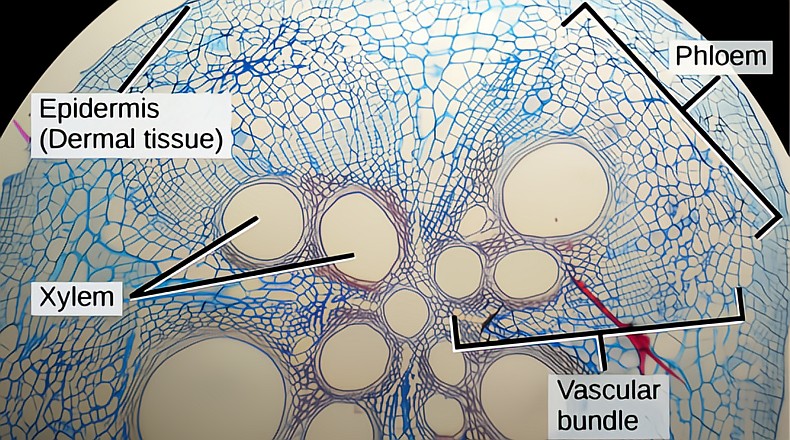
Similar in function to the xylem, the phloem is the other major vascular tissue. The phloem is responsible for the translocation of sugars produced in leaves, a very important part of the hidden life of trees. The leaves (sources) produce the products of photosynthesis, sugars which are loaded into the adjacent phloem tube cells and distributed wherever there is a demand for energy, sometimes up the stem, sometimes down and in trunks sometimes across, radially into hungry tissue. Wherever energy is needed for growth, defence, or storage, is a sink and a healthy tree’s parts will receive all that it needs. Sinks can change as the growing season progresses; common sinks are the growing tips of the SAM and RAM, the flowers, fruit, and constantly to the roots to fuel their endless work. Another common sink is storage; excess sugars are stored as starch in many tissues.
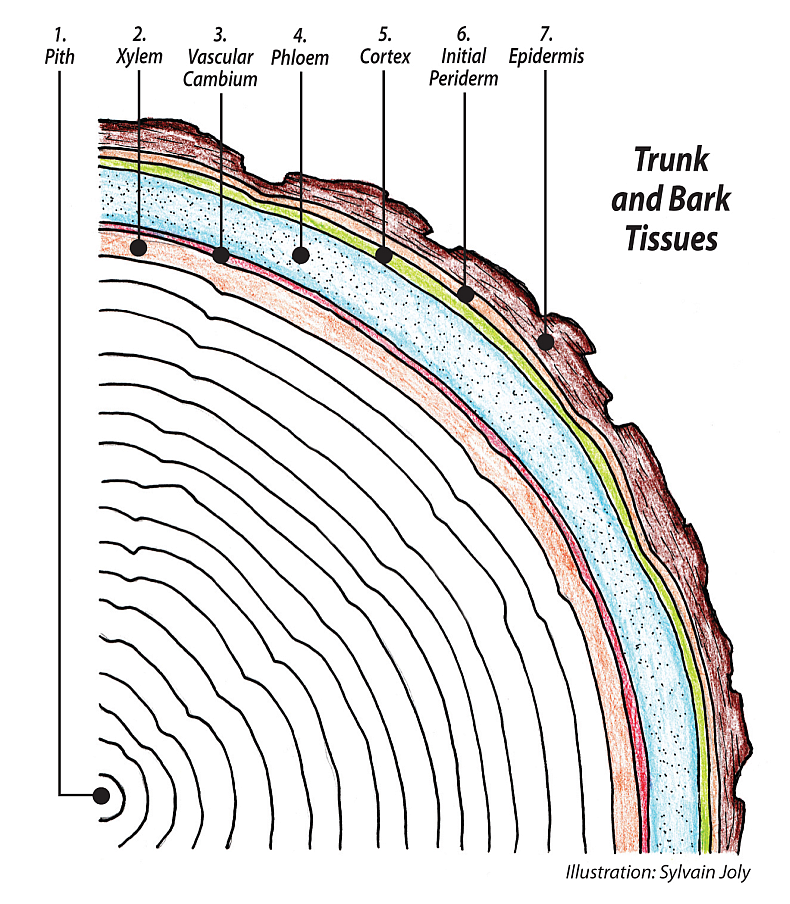
The phloem lies just to the outside of the vascular cambium, VC, and all phloem cells, sieve tube elements, STE, companion cells, CC, and ray parenchyma and others are all the daughters of the phloem fusiform and ray initials of the VC. STE are virtually hollow at maturity with most of their inner cellular components removed. This is another form of programmed cell death, which assists the overall efficiency of the tree. STE are always accompanied by a companion cell, which remains entire and is very active in the working of the phloem.
Unlike mature dead xylem tissue which can function for over a century, phloem cells are alive and usually only live for one growing season or sometimes just days or weeks. New phloem cells are constantly being replaced by the VC. This is because of the seasonal expansion of the trunk powered by the ongoing growth and increase of the xylem. The phloem cells are caught between a rock and a hard place, the expanding xylem on the inside and the stiff hard bark on the outside. A portion of all bark tissues is crushed old dead phloem cells. The phloem throughout its working season remains part of the symplast, the vast system of interconnected living tissue within the tree’s greater network. Many different signalling chemicals and hormones pass through the phloem which apart from sugar transport functions as a huge communication highway.
The phloem is important, really it is the tree’s controller, carefully moving the money (sugar) where and when it is needed and judiciously saving all the rest as starch. Dead tissue such as xylem cannot function at this level of complexity. Phloem sap is so valuable that when STE are broken or fed upon, it has a backup response that plugs the holes. P protein (phloem proteins) filaments are attached to the inside of the STE. When the STEs are damaged, P proteins are released and plug the sieve pores creating a slime plug and stopping the loss of the energy rich sap. Xylem tissue has no similar quick response.
The Munch pressure flow hypothesis proposes that sugars are loaded into the phloem tubes at sources, causing a strong sugar solution to build up; this draws water from the rays and surrounding vascular tissue. The increased pressure pushes the sugar solution in that STE through the sieve pores into the next STE. The entire phloem element becomes pressurized. At the sink the sugars are unloaded from the STE, water moves out of the cells into adjoining vascular tissue, being recycled and restoring normal pressure. Normally a cell receiving such an input of water would greatly increase its turgor pressure; not so with the STE, although alive it contains no vacuole, the organelle usually responsible for internal cell pressure. This is due to a partial programmed cell death that removes most of the inner bulky organelles yet retaining enough of them to keep it alive. With most of the cell’s inner components now missing, when the sugar concentration rises and draws water in from surrounding tissue, the water pressure pushes the sugar solution through the perforated end plate and into the next STE on its way to a sink.
Angiosperm phloem includes four cell types, gymnosperms have two. Angiosperm phloem sap moves in STEs. Single STE (cells) meet end to end to form an ST which travels from source to sink, leaves to roots. Companion cells, CC, and phloem parenchyma, PP, provide support. STE, CC and PP are all parenchyma cells. Strength is provided by phloem fibers usually positioned on the outside of the other phloem cells. Gymnosperm phloem has sieve cells, SC, and parenchyma and albuminous cells, which provide the same service that CC cells do in angiosperms, phloem loading/ unloading. Sieve cell, SC, is used instead of the angiosperm STE because gymnosperm SC are joined side to side along their tapered ends, where pit pairs meet, the cell perforations that allow fluid flow. SC do not form into long tubes. This is very similar in form to the gymnosperm tracheid, the vessel cells functioning in their xylem on the inside of the VC. Gymnosperm do not form phloem fibers; they are strong enough without them.
Tree tissue and organs are arranged predominantly along two axes. vertical and horizontal, or axial and radial. Water and nutrients need to flow in both directions. STE and CC run in the axial and phloem rays and parenchyma run in the radial direction, which ultimately feeds the cork cambium and periderm. STE in angiosperms and SC in gymnosperms are living cells responsible for translocation, the movement of sugars and other molecules from sources (leaves) to sinks (roots). The angiosperm STE is similar to a vessel in the xylem but unlike the dead at maturity vessel the STE is a living cell. With some of the inner cellular components removed, especially the vacuole, the cell is now mostly open for liquid transport. CC are intact living companion cells to each STE. STE have two types of openings to allow the movement of phloem sap, sieve plates in the end walls of angiosperms and sieve areas in the cell sidewalls. Both contain numerous sieve pores. Gymnosperms have only sieve areas. These are also called pit pairs because they line up with the sieve areas of adjoining sieve cells. Sieve pores are modified plasmodesmata. Plasmodesmata are the natural tiny openings in cell walls. They measure in nanometers and number in the thousands per cell. Plasmodesmata are the key structures to the concept of the connected network of living cells throughout the tree, the symplast.
Phloem parenchyma cells are active in radial translocation, water movement between the xylem and phloem and storage of both water and sugars. Axial phloem parenchyma cells run in a vertical direction and make up the STE, CC, phloem fiber cells and ground tissue parenchyma.
Ray parenchyma cells run in the radial direction and are called phloem rays. They are formed to the outside of the VC by the ray initials, as are the STE, which are also the daughters of the fusiform initials on the outside, (phloem side) of the VC. Water moves up in the axial xylem from the roots, enters a xylem ray (inside the VC) and is drawn horizontally into the phloem ray on the other (outer) side of the VC. Water and sugars picked up from the phloem move on outward (horizontally) and nourish the cork cambium. There are no STE in the radial system and most fluid movement is through the symplastic system of the ray parenchyma cells.
Starches, tannins and oils are some of the secondary compounds stored in phloem parenchyma, along with water in both the axial and radial tissues. Phloem parenchyma are also used for starch storage and in the fall store plenty of it, which can be the energy sources for winter metabolic activity and also to kick start the VC next spring. Tannins are used by trees for anti-herbivory and its presence in the phloem parenchyma helps keep animals out of this rich energy source.
Phloem parenchyma that stores water can act as a major back-up system when drought threatens and will supply water to the xylem, through the rays, when required.
The entire vascular system xylem/phloem has five major tissues, axial xylem, xylem rays, axial phloem, phloem rays, and a network of interwoven ground tissue made up of the xylem and phloem parenchyma, and fibers, all derived from either the outside (phloem side) or the inside (xylem side) of the vascular cambium. The various parenchyma cells interconnect the other four vascular tissues, and play a role in whole tree communication/coordination of resource allocation.
(Photo credit: “Figure 30 01 02f.jpg" by CNX OpenStax, licensed under Attribution 4.0 International (CC BY 4.0). AI upscaled, cropped, downscaled)