Roots form one of three essential organs of all plants, the other two being the stem and leaves. These organs allow plants to thrive, and plant diversity worldwide is a plain statement to their success. The roots give us another look into the hidden life of trees.
Their primary functions are absorptive anchorage, support and the uptake of minerals and water. The roots hold the stem and leaves in place so that their primary functions, energy capture (leaves) and support and fluid conduction (stems) can be as efficient as possible.
Roots also play a major role in energy storage and hormone production. Different from their up-reaching stems that have nodes for growth, and produce stems, leaves, flowers and fruit, roots only produce more roots. Each tiny extending root tip is protected by its root cap as it grows forward through the soil, seeking water and nutrients. New side (lateral) roots originate from the interior of the existing root behind the growing tip, unlike shoots which always form new tissue right at the growing tip. Root growth is controlled by nutrition, water, and gravity.
Water and ions (minerals) from the soil can move into and through the root system in two different ways. The two types of water movement are symplastic (through living cells) and apoplastic (around living cells). Symplastic movement is through tiny openings in cell walls called plasmodesmata. The fluid movement is through the root hairs into the cortex, through the endodermis and into the vascular tissue of the stele. From the stele fluid moves upward to the leaves. With the apoplastic pathway water and ions are drawn from the root hairs and through the intercellular spaces of the cortex, then through the endodermis and into the stele.
Roots are selective about what kind of molecules enter into the tree’s system. In the symplastic model it is ultimately the root hair that is allowed to make the “decision” about what enters. In the apoplastic model, a separate tissue, the casparian strip located in the cell wall spaces of the endodermis, makes the “decision.”
Most trees possess a tap root system or a modified tap root system in contrast to the fibrous root systems of grass-like plants. The tap root system is one where the primary root, the radical from the seed, grows downward and develops smaller lateral roots. Tap root systems work well in dry soils and grow towards sources of water and nitrogen and other ions in the soil.
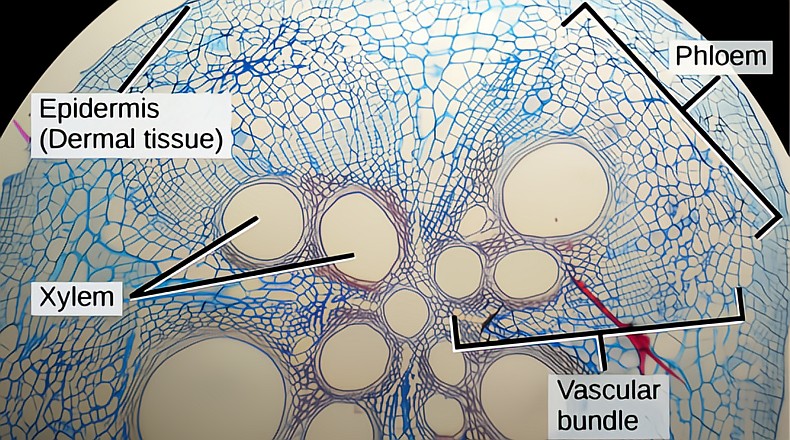
Like the shoots above, roots experience primary growth (extensions) and the RAM, root apical meristem, produces three separate differentiating meristems that combined grow all the new tissues of the root. The xylem and phloem tissues in the stele are produced by the procambium. The rhizodermis (root skin) is produced by the protoderm. The tissues of the cortex are produced by the ground meristem. All three of the tissue generating meristems are initially produced in the quiescent center, the RAM, located behind the root tip and its cap.
As the root grows we differentiate three tissue zones from youngest to oldest, the zone of division, the zone of elongation and the zone of maturation. In the division zone all the new cells for the new root and its cap are produced. It is the daughter cells produced in the zone of division that form into the three other necessary meristematic tissues, the procambium, the ground meristem and the protoderm. Further back toward the stem, and somewhat older, is the zone of elongation, where cells are growing into their mature size. These still growing cells don’t yet have any secondary walls and are not yet functioning in absorption. Further back and again somewhat older is the zone of maturation where tissue differentiation occurs. Here the vascular tissues of the stele, the xylem and phloem, the cortex and the rhizodermis are all achieving their mature form, placement and function. The stele is formed once the xylem, phloem and the endodermis (inner skin) tissues are mature.
As the new root hairs grow they fulfill their function of uptake and transport of ions and water. Root hairs appear only after elongation is finished; if not, they would be broken off from the surface of the root as it pushes through the abrasive soil particles .Many tiny root hairs, approximately 50 microns in diameter, vastly increase the root system’s overall surface area and its ability to interact with the soil.
As the root tip continually moves forward through the soil it is protected by its root cap. A coating of parenchyma cells, the root cap cells manufacture a slime called mucigel, which functions as a protective lubricant for the extending root, and its twisted path through hard soil and rocks.
A strong physical cue sensed by roots is gravity; gravitropism is the term used for plants sensing and responding to the earth’s gravity. Positive gravitropism means growing downward with the force of gravity. The radicle, the root section of the embryo in the seed, will always grow down, no matter how the seed is positioned in the soil.
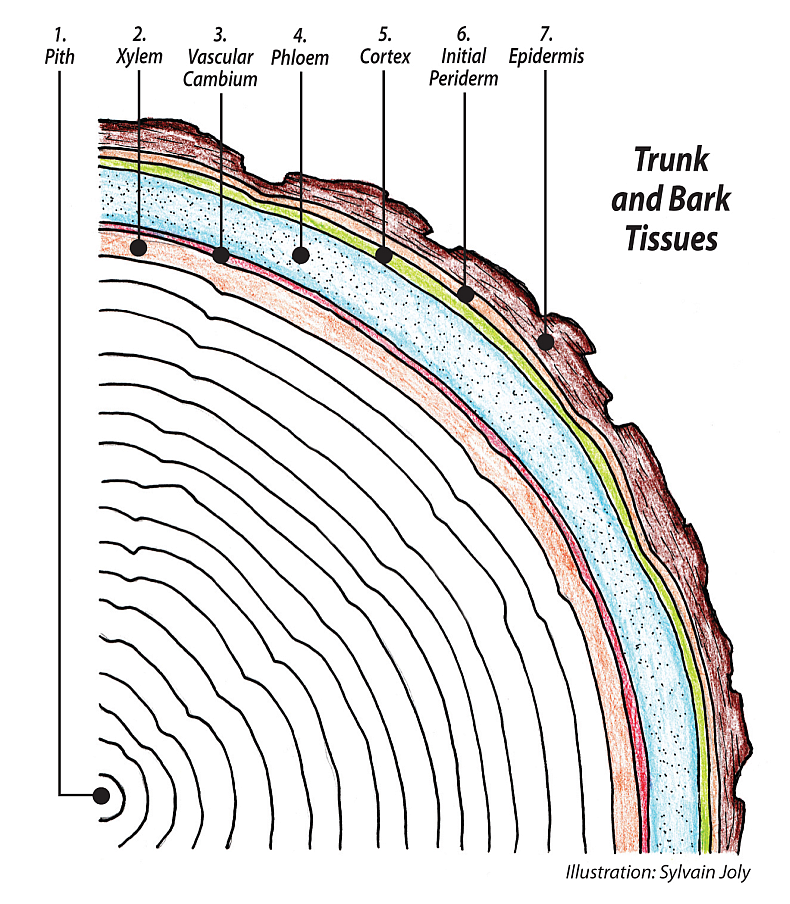
The root’s rhizodermis is the equivalent of the epidermis of the young shoots and leaves, with several distinct differences. The rhizodermis has no stomata, cuticle, produces mucigel and is specialized in water and ion absorption. It is also covered by short lived root hairs. The rhizodermis is not root bark; it is primary growth arising from the protoderm meristematic tissue. Later in its second year’s growth and ever after the root produces a periderm from the phellogen, the cork cambium; this is root bark. A periderm is the name for complete bark tissue and includes cork to the outside, the phellogen in the middle and the phelloderm to the inside right beside the cortex. Secondary growth, expansion in the root’s circumference, will strengthen the root and its ability to anchor the tree in the soil.
The zone of maturation which generates the root hairs is also the area where the newly formed stele is the most permeable. Endodermis, xylem, and phloem mature at the same rate, giving water and ions the least resistance to absorption by the xylem stream.
The ground meristem produces the cortex; its primary role is storage of water and starch. It’s much larger in volume than the cortex in stems. The roots cortex has a rhizodermis as its outside boundary and has the endodermis, inside skin that wraps the stele, as its inside boundary. The endodermis is where the casparian strip is located, acting as a filter and helping to “choose” what enters the xylem stream.
The stele is the inner vascular cylinder containing the xylem, phloem and the pericycle, bound by the endodermis to the outside. The stele contains a meristematic tissue called the pericycle which is the origin of all the lateral roots. Lateral roots are initiated in the pericycle without any involvement from the RAM. The lateral roots’ primary growth penetrates through the endodermis, the cortex and the rhizodermis tissues in order to leave the pericycle in the stele and find its way out into the soil. The tissues first stretch and then break to allow the root hair passage outside. The new root cap develops at about half way out through the cortex, and is fully functional upon emergence. The damage from the exit hole is plugged by corky cells which surround the newly emerged lateral root.
Tissue inside the new lateral root develops into phloem and xylem tubes, which connect to the parent root’s vascular tubes in the stele. It is life in the soil medium that has determined that the lateral roots do not originate from the RAM. If they did, they would all be torn off the sides as the root continues to elongate through the soil. Arising as described from the pericycle the lateral roots emerge from the maturation zone, which has stopped elongating, and the new tender lateral roots can begin their absorption work.
In their first year’s growth non-woody roots and their uncountable short-lived root hairs do almost all of the absorption work for the tree. This is the same throughout the year and every year, as new primary growth is generated by the RAM. Root hairs flourish on new lateral root growth, and sometimes only last for days. As the growing season heats up and water demands increase, so it is with root growth. The endless exploration of the soil by new roots for water and ions keeps pace with the growth and water demands of the leaves, above in the crown.
As we move into the second year of a root’s life, the growing tips and the RAM continue primary growth, as described above. The increase in diameter in the second year is called secondary growth and generates the bark of the root, the sturdy, protective layer that will cover the root during its life. Secondary growth in the year plus old root involves the formation of two additional meristems, a vascular cambium and a phellogen or cork cambium. The vascular cambium will manufacture the new secondary xylem and phloem tissues and the phellogen provides the corky periderm replacing the rhizodermis. These new meristematic tissues form perfect layers on the inside of the protophloem and are converted into cambial initials which produce xylem to the inside and phloem to the outside. Then pericycle cells outside of the protoxylem divide to grow the new cells of the vascular cambium. After some time the two new groups of dividing cells grow together to form the circular meristem we see in year-plus aged root xylem tissue.
The vascular cambium will form from the inside cells of the new meristem and the phellogen will form from the outside cells. A periderm with cork cells forms to the outside and the phellogen cork cambium to the inside. Both the vascular cambium and the cork cambium divide periclinally, adding new cells to the outside, and anticlinally, across or greater in circumference. Altogether this provides the thickening, the increase in diameter of the root as years go by. All of the initial tissues of primary growth outside the pericycle, the endodermis, the cortex, and the rhizodermis, all die and are removed. Each year the new phellogen produces the periderm for the rest of the root’s life.
As a new growth year starts, and the vascular cambium is activated, new xylem and phloem tissues increase the diameter of the root. As years of xylem tissue accumulate the root becomes larger and woody and contains growth rings similar to the trunk above.
Another interesting aspect of the life of roots is their symbiotic relationship with mycorrhizal fungus; over 5000 species of fungus form mutually beneficial relationships with trees world wide. The microscopic fungal hyphae grow round and sometimes into the non-woody young roots. This greatly increases the roots’ working surface area and penetration into the soil. The mycorrhizae greatly increase the roots’ ability to take up several essential elements and the tree is more than happy to exchange these ions for sugars produced in photosynthesis tissue elsewhere. In mutually beneficial groups of trees these mycorrhizal relationships provide another form of connection to assist the whole grove. Mics, as they are known, are so complex that they will be covered in their own essay.
(Photo credit: “Cardamine hirsuta () Photo: F.D.Richards, SE Michigan, 4/2021 - Hairy Bittercress" by F.D. Richards, licensed under CC BY-SA 2.0.)