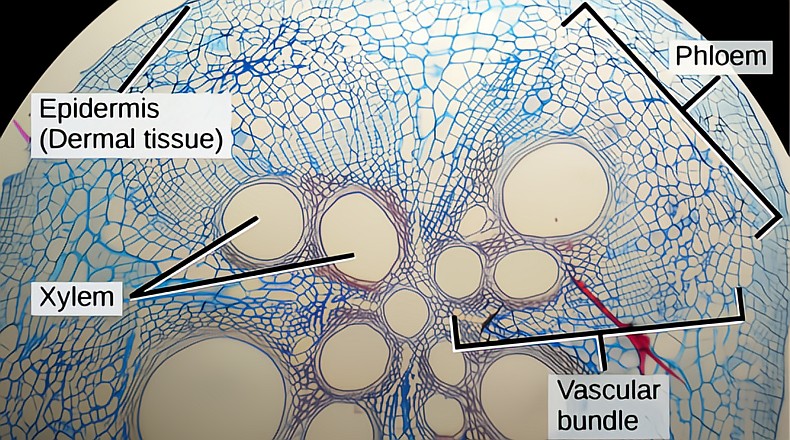
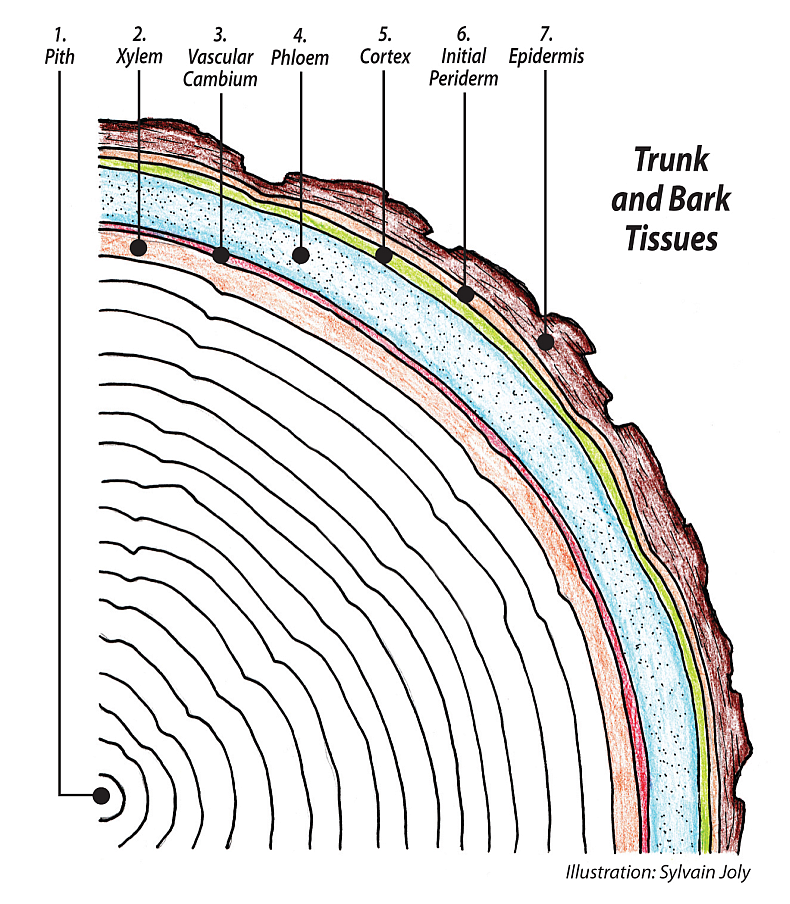
Trees are all about xylem; the bulk of the trunk is almost all xylem. There are other just as essential tissues, especially the phloem and the bark, and they both will get their own essay, but here we will concentrate on the xylem, of which 100% is generated by the vascular cambium.
The vascular cambium is the miracle bifacial meristematic tissue that grows the xylem to the inside and the phloem and bark to the outside. The cambium wraps all around the trunk and branches, everywhere under the bark, and fits like a tight suit of long johns. And that’s where it stays living, right between the xylem and the phloem for the whole life of the tree, generating new tissue on both sides, growing itself into the future.
Xylem in tree trunks is different from the xylem in leaves and the youngest roots and shoots. Xylem in trunks is not only the water conducting tracheary element, but also forms the bulk of the trunk, the wood. Buying a two-by-four from the home store is really buying a piece of xylem. In leaves and primary growth of roots and shoots, xylem and phloem piping run parallel through the tissue together, but in tree trunks and branches and roots, the two distinct vascular tissues are separated by the vascular cambium, generating the phloem to the outside and the xylem to the inside.
Xylem is the vascular tissue that allows water movement from the soil, through the roots, trunk, branches, and finally, out through the leaves. This is the process of transpiration. The evolution of a complex vasculature was essential in order for plants to become the dominant living landforms. Early plants, 450 million years ago, the bryophytes, still remain small, low to the ground and always near a water source.
Xylem is a complex tissue, made up of multiple cell types. Xylem cells can be divided into two main groups depending on their function: support in the trunk or conduction, and imperforate or perforate. Only perforate xylem cells are able to perform water conduction; all others are support.
Angiosperm (broadleaf) xylem cells include tracheids, fiber tracheids and libriform fibres, which are all imperforate, without a perforation end plate. The vessel elements are perforate and contain a perforation end wall plate, allowing water conduction. The other cell types included in the xylem are rays and parenchyma, used for storage and synthesis.
Gymnosperm (conifer) xylem cells include tracheids, parenchyma and rays. Gymnosperm tracheids are imperforate, but possess many circular bordered pits which facilitate efficient water movement.
Water movement, whether in soils, roots, trunks, branches, leaves, or the atmosphere is always controlled by the water concentration. Water always moves from a wetter, higher concentration area to a dryer, lower concentration area. Dry soil near wet soil draws water to become wetter. Roots needing water and having a lower water concentration than well-watered soil draw water from the soil. The trunk xylem tissues, having a lower water concentration than the roots, draw water from the roots, etc., etc. and on up we go. The driest, lowest water concentration is almost always the atmosphere. It is during daylight when leaf stomata are open and losing a lot of water (transpiration) that water movement from the soil, through the tree and into the atmosphere is at its highest.
Heavy water requirements are a bit of a design fault in trees. There just wasn't any other option for conducting photosynthesis, having thousands of tiny leaf stomata open to allow in the essential CO2 required. At least 95% of water drawn up into trunks and passed to the leaves evaporates out from the stomata. In nature, trees are only found where their minimum water requirements are met, rarely in grasslands and deserts.
This is a sketch of a process called the cohesion/tension theory, or the ascent of sap. Water loves water, has a high surface tension, is sticky with itself, and is highly cohesive (hard to pull apart). It can also stand a very high tension; in some trees, a water column is drawn upwards hundreds of feet. The evaporation from the leaves draws a tiny water column up through each functioning vessel element, connected all the way down to the roots. A vessel can be dozens or even hundreds of individual vessel cells connected end to end. Tracheids don't do this. In angiosperms, the perforate vessels accomplish this with water movement through perforation end plates at the ends of each cell. Gymnosperms accomplish the same hydraulics by the use of pits, thousands of tiny openings in the side walls of their tracheids.
Occasionally, in the highly stressful conditions of extreme heat and or drought, the amount of evaporation from the leaves outstrips all of the roots' ability to meet their demands. When this happens, when too much tension is applied to the water column in the vessel/tracheid, it will break, or cavitate. This is similar to an embolism; the vessel element will fill with water vapour and flow through that vessel element will cease. Two backup functions are in place to deal with this—one is hydraulic and the other is mechanical.
The functioning vessel elements are surrounded by living parenchyma cells, which can contribute enough water to refill the cavitated vessel element, thus restoring the water column. The second mechanical repair involves an inner membrane and a tiny blocking cell; there are the margo and torus that function at cavitated pit pairs to restore the water column.
Water flow is related to vessel element diameter. The wider the vessel element, the greater the flow. As mentioned, water is sticky and binds to itself and the inside surface of the vessel element (cohesion). This bound water reduces the free flow in the vessel element, so wider vessel elements move the most water but are the most susceptible to cavitation. This is the conflict in vessel element design. Most trees adapt to this by varying their vessel element diameters in different times of the growing season. Water is usually abundant in the spring, hence early wood usually has greater diameter vessel elements than late wood vessel elements, due to failing water availability as the summer progresses.
Tracheids are imperforate vessel elements and the only water conductors in gymnosperms. Tracheids are long and thin with tapered ends. Water movement upwards is from tracheid to tracheid via the numerous pit pairs in the tracheid wall. The pits begin as plasmodesmata and are formed between spaces in the inner structural reinforcements of the secondary inner wall. Tracheids vary in length from 100 to 6,000 microns—that’s 0.1 mm up to a whopping 6 mm—and from 10 to 70 microns in width. Average tracheid length in gymnosperms is 3,500 microns, and 1,200 microns in angiosperms. Gymnosperm tracheids are more primitive in design than vessels and show a greater variety of size and shape than angiosperm tracheids and vessels.
Vessels give the angiosperms greater adaptive resistance and allow them to occupy many diverse biomes that do not favour gymnosperms. The evolutionary development was from tracheids to fibre tracheids to libriform fibres to vessel elements. During cell development, the required pattern of secondary wall reinforcement forms to the sides and on the perforation plate on the inside of the primary cell wall. At maturity, the initial cell wall is degraded and removed, allowing the pits to function and align with adjacent pits in other vessel elements. The vessel element is now dead but fully functional; indeed, more so than the developing vessel element cell which, when full of its protoplasm and organelles, would seriously impede water transport through it.
The five patterns of vessel element inner cell wall thickening are, in increasing strength, complexity and evolutionary age, annular, helical, scaliform, reticulate and nearly continuous. Annular are simple rings of secondary wall material. Helical is secondary wall material that wraps around inside the primary wall. Only these two lighter forms (collenchyma) of reinforcement are used in growing, still expanding cells, allowing them to continue to stretch. Scaliform thickening runs across the inside of the inner cell wall and contains about twice as much reinforcement as annular provides. Reticulate thickening is irregular and net-like; cells with recticulate inner wall construction are stiff and strong. Cells with continuous inner reinforcement have nearly solid secondary wall construction with openings only for pits, pit pairs and end wall perforation plates. These last three types (schlerenchyma) occur only after cell growth and expansion is complete. They are too strong and are only applied after the growing cell is complete.