Leaves are tirelessly active as much as 16 hours a day in the height of the growing season. Although they seem soft, they need to be flexible; they are tough and endure cold, hail, snow, hard rain, strong winds and insect attacks. These are just some of their problems, all the while producing sugars which will supply the energy for the whole year of the tree’s life. Given the most basic set of tools, sunlight, air, water, and minerals from the soil, trees grow their entire body, producing all of the materials needed for their lives, sugars, amino acids, proteins, lipids, nucleic acids, vitamins, everything they need, and with no waste. Leaves truly are a hidden window into the life of trees.
The usual broadleaf leaf blade, the lamina, is flat with two sides; it is bifacial. The upper towards the light is the adaxial side, the lower underside facing the earth is the abaxial side. The lamina is attached to the twig by the petiole, which connects the leaf to the tree’s vascular system, the xylem and phloem. Conifers have long-lived flat elongated needle-like leaves that are excellent at water conservation.
Leaves are very thirsty due to their constant water loss. During the process of photosynthesis the stomata, the leaf pores, need to be wide open to allow the free inflow of CO2 from the atmosphere. While open, a tremendous amount of water vapor transpires from the exposed inner leaf surface, the spongy mesophyll. A hundred gallons a day for a large tree is not an exaggeration. The essential function of water in leaves is twofold; it is a necessary component in the chemical reaction of photosynthesis and is also paramount in cooling the leaf, always in the sun, as it evaporates away. The necessary water is delivered to each leaf by the tree’s vascular system, which spreads out and divides throughout the leaf via its venation system.
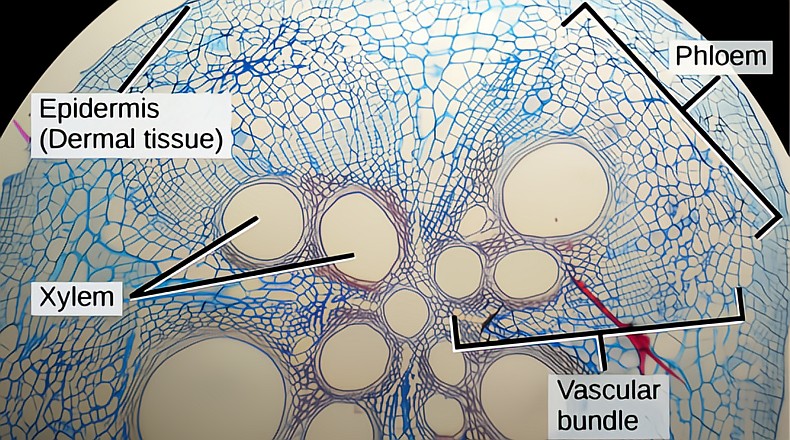
The veins contain the two separate pipes of the vascular system, the xylem, bringing water and mineral ions and the phloem receiving the produced sugars. These two separate pipes are side by side in a bundle. The splitting and spreading from larger to smaller veins that make their pattern throughout the leaf tissue is called reticulate venation. There are two distinct vein sizes, major and minor; the majors are the main conduits and also supply physical support due to their reinforced cell walls. The major vein of the leaf’s center which connects to the petiole is called the midrib. The job of the minor veins is to spread the water and ions of the xylem stream throughout the mesophyll and to load the phloem pipes, sieve tube elements, with the sugars produced in the chloroplasts of the mesophyll.
No inner leaf cell is more than a few cells away from a minor vein. A minor vein contains a tiny vascular bundle, a few phloem sieve tube elements and a few xylem elements inside the bundle sheath. There are two main types of minor leaf veins, open veins with symplastic connections via the cell wall pores, the plasmodesmata, and closed veins with apoplastic connections, connections with the spaces outside and between cells.
The form of leaves is optimized for water retention, gas exchange and light absorption. Foliage leaves’ primary function is photosynthesis. The leaves’ epidermis consists of guard cells, trichomes, leaf hairs, pavement cells and the outer waxy cuticle. Below the leaves’ upper surface are the rows of vertically aligned palisade parenchyma cells, chloroplasts. Below those closer to the underside of the leaf is the majority of the inner spongy mesophyll, which has easy access to the inner leaf spaces, allowing a free movement of CO2 gas that has entered from the stomata. Nowhere else in the tree’s body is there such a high concentration of chlorophyll-containing chloroplasts cells in which photosynthesis occurs. Inside the lower portion of the leaf the intercellular air spaces allow half of the chloroplast cell surface to be in contact with CO2 gas. This is where the term spongy mesophyll comes from. The remaining cell to cell surfaces have plasmodesmata in their millions, allowing for symplastic efficiency between the mesophyll and the vascular system.
Differing amounts of light penetrate through the tree during leaf development and that has some impact on the leaf’s final form. When light availability is low, especially inside the crown, shade leaves are formed. Usually larger than sun leaves that mature in the light-rich edges of the crown, shade leaves are also thinner, allowing the light they do receive to penetrate fully into the leaf to be as photosynthetically efficient as possible. Shade leaves have a single layer of palisade cells, less interior air space and a thinner cuticle, and also a greater percentage of chlorophyll than sun leaves. Sun leaves can be five times as photosynthetically productive as shade leaves.
Conifer leaves are quite different. Every aspect of conifer needle design speaks of their overall high water-conserving ability. Evolved for the north and mountainous country, conifers need to be able to deal with a lot of snow, wet conditions in the spring and later drought conditions on the dry mountain slopes they survive on. Mountain country receives extra strong doses of sunlight, good for photosynthesis but also highly evaporative.
Conifers retain their needles for many years, sometimes as much as 30 years. Larches are one of the few examples within the large group of conifers that grow and then drop their needles each year. A trademark of almost all conifers is xeromorphy, the natural adaptation to dry conditions, which is accomplished by outer pavement cells with secondary cell walls and a thick waxy cuticle. They also have deeply recessed stomata. The naturally economical form of conifer needles reduces the surface to volume ratio, which helps reduce transpiration.
Northern broadleaf trees have evolved to bear leaves only during the warmer half of the year, when they can survive. Sometimes our trees bear leaves for less than six months, but remain alive and functioning through the repeated twelve-month cycles. This strongly says something about the value of leaves and the energy they produce, being able to generate enough energy in six months to last through twelve. The sugars are converted to starch and then stored in tissues from the roots to the branches, and spent as needed. Trees have lots of ongoing costs, expenses, just like all living things, the metabolic cost of being alive, the cost of growth and defence. It all adds up. Each expenditure takes away from the year’s income. Trees are naturally balanced living systems with three main organs, leaves, trunk, roots, all in sync, all balanced. There are no “extra” leaves; they don’t need pruning, except common-sense pruning.
The tree service industry ignores these basic facts. Most arborists have a much too ultra-clean vision for your trees, and serious overpruning and thinning is how that look is accomplished. Have you ever had your trees pruned, only to have them react with a lot of sucker growth, either that same year or the next? This is the surest sign of heavy over pruning; the tree is screaming, OVERPRUNING. Arborists need to shift their attention from one of aesthetics to one of energy production/flow/usage. If they did, the trees would be a lot healthier. I honestly feel the health of the tree is more valuable than an overpruned pretty look.
This is not to say that from time to time some pruning need not be done. Always remove the 3 Ds, diseased, dead and damaged. House, sidewalk and driveway clearances are just being practical. But go light in the rest of the crown. We are not smarter than trees about how to live their lives. Interview your arborist and look for empathy towards your trees. An arborist trained as a removal logger who just loves to cut wood is not the person to do a careful job on your apple tree.
Our broadleaf trees are winter deciduous, evolved to shed their leaves as the signs of approaching winter appear. The process is called autumnal senescence. Before the leaf is actually shed from the tree a process of resource extraction occurs. Valuable molecules of proteins, amino acids, and many mineral ions concentrated in chloroplasts need to be recovered. The seasonal change clues of shorter days and colder nights are the signals that begin the process. If all goes well, all the proteins, amino acids and ions will be recovered, recycled, downloaded into the phloem and pumped back into the body of the tree for storage. All of the actions of senescence are the result of inherent gene expressions.
After the breakup and recycling of the green chlorophyll-rich chloroplasts, the remaining pigments in the leaf can be seen. This amazing fall color show is beautiful in every region. The two main pigments that supply the rich yellows, oranges and reds are anthocyanins and carotenoids.
Once the colors fade it is time for abscission or leaf drop, the safe removal of the leaves from the twigs. Leaf abscission starts with the formation of the abscission zone, which forms at the base of the petiole. During the process a separation layer of corky cells is formed, which later allows the petiole to break off without tearing any of the twig tissue. What is left behind on the twig is the leaf scar with the major vein sites clearly seen.